El segundo gran modelo de organización celular es el característico de los organismos eucariotas, que incluyen a los protistas, los hongos, las plantas y los animales. Aunque existen diferencias significativas entre ellos, todos comparten la misma organización básica, caracterizada por la compartimentalización interna, posible gracias a la presencia de un sistema de membranas intracelular.
La compartimentalización ofrece a la célula nuevas posibilidades; por una parte, los diferentes compartimentos pueden especializarse en funciones distintas, lo que facilita la coordinación entre distintas funciones celulares y la realización de varias al mismo tiempo. Por otra parte, las membranas intracelulares son utilizadas como mecanismo de producción de energía mediante la creación de gradientes quimiosmóticos en el interior de la célula (mitocondria, cloroplasto). Por último, los diferentes compartimentos pueden tener ambientes químicos distintos, incluso totalmente diferentes. Por ejemplo, el citoplasma de las células eucariotas es anaerobio, hasta el punto de que el oxígeno resulta tóxico en esa zona de la célula, mientras que la matriz mitocondrial es aerobia, y permite la utilización del oxígeno en procesos metabólicos.
De hecho, se supone que el origen de las células eucariotas es el resultado de la necesidad de adaptarse a un ambiente químico diferente al que existía previamente. Según la hipótesis endosimibionte, antes de la revolución del oxígeno (la aparición de la fotosíntesis oxigénica que dio lugar a que la atmósfera acumulara oxígeno) solo existían células procariotas. La acumulación de oxígeno en la atmósfera propició la aparición de algunas células también procariotas capaces de utilizarlo en sus procesos metabólicos (las proto-mitocondrias), mientras que otras habrían evolucionado "englobando" con su membrana este tipo celular (y dando lugar a los eucariotas heterótrofos) o también a las células fotosintéticas (proto-cloroplastos), originando los eucariotas autótrofos. El sistema endomembranoso (los orgánulos como el retículo endoplásmico, el aparato de Golgi, las vacuolas o la membrana nuclear) habría evolucionado previamente a partir de invaginaciones de la membrana plasmática de la célula.
Aunque no existe una "célula eucariota generalizada" es posible hacer un listado de todos los orgánulos y estructuras comunes a todas las células con este grado de organización, si bien hay que tener en cuenta que las células reales difieren siempre de este modelo simplificado.

Las características generales de este tipo de células son las siguientes:
El material genético de los procariotas es mucho más complejo que el de los procariotas. Consta siempre de varias moléculas, lo que supone una cantidad de información considerablemente mayor. Además, el ADN está organizado en forma de cromosomas, estructuras en las que un conjunto de proteínas (entre ellas las histonas) empaquetan y protegen el material genético propiamente dicho. Por último, el material genético y todos los compuestos relacionados directamente con su replicación y expresión están separados del resto del citoplasma por un sistema membranoso.
La organización del citoplasma también es mucho más compleja: tiene una estructura de sostén, constituida por varios tipos de proteínas, que recibe el nombre de citoesqueleto. Además posee varios sistemas de membrana, diferenciados en estructura y función (orgánulos membranosos).
Orgánulos comunes a todas las células eucariotas
Las células eucariotas incluyen dos tipos diferentes de ribosomas: los de su citoplasma tienen un coeficiente de sedimentación (parámetro relacionado con su masa y con su forma) de 80S. Su subunidad mayor tiene un coeficiente de sedimentación de 60S y la pequeña de 40S. Este tipo de ribosomas se encuentran tanto libres en el citoplasma como adheridos a algunos sistemas de membrana del citoplasma, concretamente a una parte de su retículo endoplásmico. Los ribosomas libres se encargan de sintetizar las proteínas que la célula necesita en su citoplasma, mientras que los que están unidos al retículo se encargan de sintetizar proteínas que van a quedar en el interior del sistema endomembranoso (en el aparato de Golgi, en el retículo endoplásmico o en algunas vesículas especializadas, como los lisosomas) o que van a ser secretadas al exterior de la célula para que cumplan su función allí.
Pero, por otra parte, las células eucariotas cuentan también con ribosomas 70S, idénticos a los que se encuentran en los procariotas, en el interior de sus orgánulos más complejos: mitocondrias y plastos. Este hecho, junto a la presencia de ADN circular y cerrado y la doble membrana de estos orgánulos, apoya la hipótesis endosimbionte para explicar su origen.

Todas las células eucariotas poseen también vesículas o bolsas de membrana, de estructura sencilla (básicamente esféricas, o deformadas), que se diferencian entre sí fundamentalmente por su función. Entre esas estructuras se encuentran las vacuolas de las células vegetales (su nombre procede de vacuum, vacío, porque al microscopio no se aprecia su contenido; en realidad contienen líquido de composición diferente al citoplasma), las vesículas de transporte que se mueven entre diferentes orgánulos celulares llevando sustancias que no se mezclan con el citoplasma, o cuerpos más o menos especializados como los lisosomas, que contienen enzimas capaces de romper otras proteínas, y que no pueden mezclarse con otros componentes celulares sin riesgo de destruirlos. Los peroxisomas se ocupan de eliminar sustancias tóxicas de la célula mediante su oxidación.
El siguiente nivel de complejidad en las estructuras intracelulares está constituido por los sistemas membranosos complejos (retículo endoplásmico liso y rugoso, aparato de Golgi y membrana celular), que pueden diferenciarse entre sí tanto por su morfología como por su función.
 El retículo endoplásmico liso es un conjunto de cisternas de membrana, típicamente en forma de tubo, que en su conjunto forman un sistema de tuberías interconectadas que recorren buena parte de la célula, aunque puede tener distintos aspectos en células diferentes. Su principal característica distintiva es que no presenta ribosomas adheridos. Las funciones que realiza son las siguientes:
El retículo endoplásmico liso es un conjunto de cisternas de membrana, típicamente en forma de tubo, que en su conjunto forman un sistema de tuberías interconectadas que recorren buena parte de la célula, aunque puede tener distintos aspectos en células diferentes. Su principal característica distintiva es que no presenta ribosomas adheridos. Las funciones que realiza son las siguientes:
 El retículo endoplásmico rugoso es un conjunto de cisternas y conductos que se distribuye por toda la célula, aunque su aspecto más típico es el de un conjunto de sacos aplanados. El término "rugoso" que se utiliza para designarlo se refiere a su aspecto en las micrografías electrónicas, en las que se observa que su membrana presenta, hacia su cara citoplasmática, puntos oscuros que sobresalen de la línea de membrana. Se trata de ribosomas que se encuentran adheridos a la superficie de la membrana y que vierten las proteínas que sintetizan directamente al interior de las vesículas manteniendolas, por tanto, separadas del citoplasma. Las funciones que realiza son:
El retículo endoplásmico rugoso es un conjunto de cisternas y conductos que se distribuye por toda la célula, aunque su aspecto más típico es el de un conjunto de sacos aplanados. El término "rugoso" que se utiliza para designarlo se refiere a su aspecto en las micrografías electrónicas, en las que se observa que su membrana presenta, hacia su cara citoplasmática, puntos oscuros que sobresalen de la línea de membrana. Se trata de ribosomas que se encuentran adheridos a la superficie de la membrana y que vierten las proteínas que sintetizan directamente al interior de las vesículas manteniendolas, por tanto, separadas del citoplasma. Las funciones que realiza son:
 El aparato de Golgi es un conjunto de sacos aplanados, que reciben el nombre de dictiosomas. Se encuentran apilados unos sobre otros, y típicamente tienen un aspecto curvado, en general con la cara convexa orientada hacia el núcleo. Sus funciones son, fundamentalmente:
El aparato de Golgi es un conjunto de sacos aplanados, que reciben el nombre de dictiosomas. Se encuentran apilados unos sobre otros, y típicamente tienen un aspecto curvado, en general con la cara convexa orientada hacia el núcleo. Sus funciones son, fundamentalmente:
El núcleo celular es una estructura que contiene el material genético celular en el interior de un sistema membranoso. Los elementos que lo forman son:

 La mitocondria es uno de los orgánulos celulares de mayor complejidad. Está presente en la práctica totalidad de las células eucariotas. Al microscopio aparece como un cuerpo alargado, del tamaño y forma de un bacilo. La observación de su estructura permite apreciar que posee un sistema de dos membranas, separadas entre sí por una zona llamada espacio intermembranoso. La membrana interna suele presentar un conjunto de invaginaciones en forma de tabique que reciben el nombre de crestas mitocondriales.
La mitocondria es uno de los orgánulos celulares de mayor complejidad. Está presente en la práctica totalidad de las células eucariotas. Al microscopio aparece como un cuerpo alargado, del tamaño y forma de un bacilo. La observación de su estructura permite apreciar que posee un sistema de dos membranas, separadas entre sí por una zona llamada espacio intermembranoso. La membrana interna suele presentar un conjunto de invaginaciones en forma de tabique que reciben el nombre de crestas mitocondriales.

La presencia de las crestas mitocondriales indica que la mitocondria maximiza la proporción entre la superficie de su membrana interna y su volumen, lo que indica que la membrana interna tiene una importancia considerable en su funcionamiento. Se sabe, en efecto, que una parte importante de los procesos metabólicos que ocurren en la mitocondria se producen, precisamente, en su membrana interna.
El interior de la mitocondria se denomina matriz mitocondrial, y es bastante similar al interior de una célula procariota. Al igual que estas células, presenta un genóforo propio, constituido por una única molécula de ADN bicatenario circular, cerrado y no asociado a histonas. Los genes de este genóforo se expresan en la mitocondria, y dan lugar a algunas (pero no a todas) de las proteínas que la mitocondria utiliza en su funcionamiento. También aparecen en la matriz mitocondrial ribosomas similares a los bacterianos, es decir, con un coeficiente de sedimentación de 70S y subunidades 50S y 30S. Estas características comunes con los procariotas refuerzan la hipótesis de su origen endosimbionte. También apoya esta hipótesis el hecho de que la mitocondria se reproduzca autónomamente dentro de la célula, replicando y transmitiendo su material genético a las nuevas mitocondrias, sin que haya relacion directa con los procesos generales de reproducción celular.
En cuanto a las funciones mitocondriales, la más importante es la de proporcionar energía al resto de la célula, gracias a que en su interior (matriz celular y membrana interna) ocurren algunas de las rutas metabólicas más importantes: el ciclo de Krebs, la fosforilación oxidativa y la β-oxidación. Puesto que la mitocondria es el único orgánulo celular capaz de utilizar el oxígeno, es en su interior donde finalizan los procesos degradativos de los combustibles metabólicos. Algunas de estas rutas metabólicas, en particular el ciclo de Krebs, sirven también para proporcionar a la célula sustancias que intervienen en diferentes procesos celulares (intermediarios metabólicos).
El citoesqueleto o esqueleto celular es un conjunto de estructuras filamentosas de naturaleza proteica entre cuyas funciones destacan mantener la forma celular, o permitir sus cambios, permitir la movilidad o inmovilidad de los orgánulos y participar en la división del citoplasma celular (citocinesis) durante los procesos de división celular.

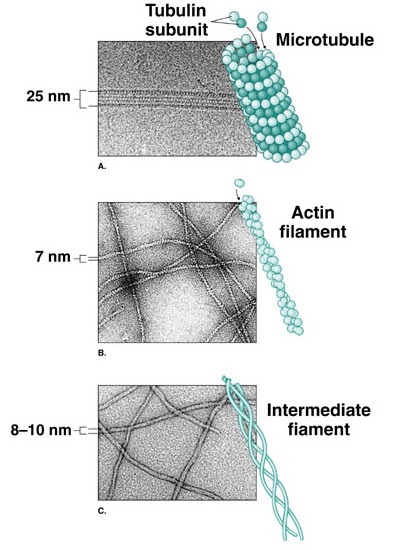 El citoesqueleto tiene tres tipos de componentes fundamentales: filamentos de actina (microfilamentos), filamentos intermedios y microtúbulos.
El citoesqueleto tiene tres tipos de componentes fundamentales: filamentos de actina (microfilamentos), filamentos intermedios y microtúbulos.
La compartimentalización ofrece a la célula nuevas posibilidades; por una parte, los diferentes compartimentos pueden especializarse en funciones distintas, lo que facilita la coordinación entre distintas funciones celulares y la realización de varias al mismo tiempo. Por otra parte, las membranas intracelulares son utilizadas como mecanismo de producción de energía mediante la creación de gradientes quimiosmóticos en el interior de la célula (mitocondria, cloroplasto). Por último, los diferentes compartimentos pueden tener ambientes químicos distintos, incluso totalmente diferentes. Por ejemplo, el citoplasma de las células eucariotas es anaerobio, hasta el punto de que el oxígeno resulta tóxico en esa zona de la célula, mientras que la matriz mitocondrial es aerobia, y permite la utilización del oxígeno en procesos metabólicos.
De hecho, se supone que el origen de las células eucariotas es el resultado de la necesidad de adaptarse a un ambiente químico diferente al que existía previamente. Según la hipótesis endosimibionte, antes de la revolución del oxígeno (la aparición de la fotosíntesis oxigénica que dio lugar a que la atmósfera acumulara oxígeno) solo existían células procariotas. La acumulación de oxígeno en la atmósfera propició la aparición de algunas células también procariotas capaces de utilizarlo en sus procesos metabólicos (las proto-mitocondrias), mientras que otras habrían evolucionado "englobando" con su membrana este tipo celular (y dando lugar a los eucariotas heterótrofos) o también a las células fotosintéticas (proto-cloroplastos), originando los eucariotas autótrofos. El sistema endomembranoso (los orgánulos como el retículo endoplásmico, el aparato de Golgi, las vacuolas o la membrana nuclear) habría evolucionado previamente a partir de invaginaciones de la membrana plasmática de la célula.
Aunque no existe una "célula eucariota generalizada" es posible hacer un listado de todos los orgánulos y estructuras comunes a todas las células con este grado de organización, si bien hay que tener en cuenta que las células reales difieren siempre de este modelo simplificado.

- Nucleoplasma:
- El material genético de la célula siempre está formado por varias moléculas.
- Los cromosomas son moléculas de ADN de cadena doble, lineal y terminadas en secuencias de ADN especiales llamadas "telómeros"
- El ADN está asociado a histonas.
- El conjunto del material genético está rodeado de una doble membrana (membrana nuclear) formando un compartimento separado, el núcleo.
- El material genético se replica mediante mitosis.
- Organización del citoplasma:
- Los ribosomas tienen un coeficiente de sedimentación de 80S (son algo más grandes que los bacterianos).
- Poseen citoesqueleto y corrientes citoplasmáticas.
- Existen orgánulos diferenciados, constituidos por membrana, con funciones especializadas.
El material genético de los procariotas es mucho más complejo que el de los procariotas. Consta siempre de varias moléculas, lo que supone una cantidad de información considerablemente mayor. Además, el ADN está organizado en forma de cromosomas, estructuras en las que un conjunto de proteínas (entre ellas las histonas) empaquetan y protegen el material genético propiamente dicho. Por último, el material genético y todos los compuestos relacionados directamente con su replicación y expresión están separados del resto del citoplasma por un sistema membranoso.
La organización del citoplasma también es mucho más compleja: tiene una estructura de sostén, constituida por varios tipos de proteínas, que recibe el nombre de citoesqueleto. Además posee varios sistemas de membrana, diferenciados en estructura y función (orgánulos membranosos).
Orgánulos comunes a todas las células eucariotas
Las células eucariotas incluyen dos tipos diferentes de ribosomas: los de su citoplasma tienen un coeficiente de sedimentación (parámetro relacionado con su masa y con su forma) de 80S. Su subunidad mayor tiene un coeficiente de sedimentación de 60S y la pequeña de 40S. Este tipo de ribosomas se encuentran tanto libres en el citoplasma como adheridos a algunos sistemas de membrana del citoplasma, concretamente a una parte de su retículo endoplásmico. Los ribosomas libres se encargan de sintetizar las proteínas que la célula necesita en su citoplasma, mientras que los que están unidos al retículo se encargan de sintetizar proteínas que van a quedar en el interior del sistema endomembranoso (en el aparato de Golgi, en el retículo endoplásmico o en algunas vesículas especializadas, como los lisosomas) o que van a ser secretadas al exterior de la célula para que cumplan su función allí.
Pero, por otra parte, las células eucariotas cuentan también con ribosomas 70S, idénticos a los que se encuentran en los procariotas, en el interior de sus orgánulos más complejos: mitocondrias y plastos. Este hecho, junto a la presencia de ADN circular y cerrado y la doble membrana de estos orgánulos, apoya la hipótesis endosimbionte para explicar su origen.
Todas las células eucariotas poseen también vesículas o bolsas de membrana, de estructura sencilla (básicamente esféricas, o deformadas), que se diferencian entre sí fundamentalmente por su función. Entre esas estructuras se encuentran las vacuolas de las células vegetales (su nombre procede de vacuum, vacío, porque al microscopio no se aprecia su contenido; en realidad contienen líquido de composición diferente al citoplasma), las vesículas de transporte que se mueven entre diferentes orgánulos celulares llevando sustancias que no se mezclan con el citoplasma, o cuerpos más o menos especializados como los lisosomas, que contienen enzimas capaces de romper otras proteínas, y que no pueden mezclarse con otros componentes celulares sin riesgo de destruirlos. Los peroxisomas se ocupan de eliminar sustancias tóxicas de la célula mediante su oxidación.
El siguiente nivel de complejidad en las estructuras intracelulares está constituido por los sistemas membranosos complejos (retículo endoplásmico liso y rugoso, aparato de Golgi y membrana celular), que pueden diferenciarse entre sí tanto por su morfología como por su función.
- Participa en la síntesis de lípidos (ácidos grasos, triacilglicéridos y fosfolípidos) y en el transporte de estas sustancias a través de la célula.
- Posee enzimas detoxificantes, que metabolizan y eliminan el alcohol y otras sustancias químicas.
- Síntesis de proteínas destinadas al interior de las vesículas de membrana (lisosomas), que se incorporan a las membranas (proteínas integrales de membrana) o destinadas a la secreción.
- Glucosilación de proteínas.
- Circulación intracelular de sustancias que no deben mezclarse con el contenido citoplasmático.
- Modificación química de diversas sustancias producidas en la célula, para que puedan adquirir su forma activa (glucosilación, fosforilación) o para identificar su destino final, dentro o fuera de la célula (se unen sustancias que actúan como señal, en particular oligosacáridos).
- Secreción celular: todas las sustancias secretadas por la célula pasan por el aparato de Golgi.
- Producción de membrana para otros sistemas celulares, como los lisosomas o la membrana externa de la célula.
El núcleo celular es una estructura que contiene el material genético celular en el interior de un sistema membranoso. Los elementos que lo forman son:
- La envoltura nuclear, una doble membrana que lo separa del citoplasma y que se continúa con el retículo endoplásmico rugoso. Al igual que éste, la cara externa de la membrana presenta ribosomas adosados. El sistema de doble membrana está atravesado por "poros", estructuras proteicas complejas que desempeñan una función de "diafragma", ya que tienen la posibilidad de cerrarse o abrirse, permitiendo la salida de moléculas de ARN.
- La lámina nuclear, una zona situada inmediatamente por debajo de la zona interna de la membrana nuclear, formada por filamentos proteicos que tienen como función proporcionar sostén al núcleo.
- El nuceloplasma es la zona poco diferenciada que contiene el resto de los elementos nucleares.
- El nucleolo es una zona que presenta aspecto diferente al resto del núcleo. Se sabe que en él se está sintetizando activamente ARN ribosómico a lo largo de casi todo el tiempo.
- La cromatina es el conjunto de material genético de la célula. Durante la interfase, es decir, mientras la célula está funcionalmente activa, se encuentra desespiralizada, por lo que no se observan los cromosomas. Por el contrario, durante los periodos de división, el ADN se empaqueta hasta formar los cromosomas, que sí pueden observarse.

La presencia de las crestas mitocondriales indica que la mitocondria maximiza la proporción entre la superficie de su membrana interna y su volumen, lo que indica que la membrana interna tiene una importancia considerable en su funcionamiento. Se sabe, en efecto, que una parte importante de los procesos metabólicos que ocurren en la mitocondria se producen, precisamente, en su membrana interna.
El interior de la mitocondria se denomina matriz mitocondrial, y es bastante similar al interior de una célula procariota. Al igual que estas células, presenta un genóforo propio, constituido por una única molécula de ADN bicatenario circular, cerrado y no asociado a histonas. Los genes de este genóforo se expresan en la mitocondria, y dan lugar a algunas (pero no a todas) de las proteínas que la mitocondria utiliza en su funcionamiento. También aparecen en la matriz mitocondrial ribosomas similares a los bacterianos, es decir, con un coeficiente de sedimentación de 70S y subunidades 50S y 30S. Estas características comunes con los procariotas refuerzan la hipótesis de su origen endosimbionte. También apoya esta hipótesis el hecho de que la mitocondria se reproduzca autónomamente dentro de la célula, replicando y transmitiendo su material genético a las nuevas mitocondrias, sin que haya relacion directa con los procesos generales de reproducción celular.
En cuanto a las funciones mitocondriales, la más importante es la de proporcionar energía al resto de la célula, gracias a que en su interior (matriz celular y membrana interna) ocurren algunas de las rutas metabólicas más importantes: el ciclo de Krebs, la fosforilación oxidativa y la β-oxidación. Puesto que la mitocondria es el único orgánulo celular capaz de utilizar el oxígeno, es en su interior donde finalizan los procesos degradativos de los combustibles metabólicos. Algunas de estas rutas metabólicas, en particular el ciclo de Krebs, sirven también para proporcionar a la célula sustancias que intervienen en diferentes procesos celulares (intermediarios metabólicos).
El citoesqueleto o esqueleto celular es un conjunto de estructuras filamentosas de naturaleza proteica entre cuyas funciones destacan mantener la forma celular, o permitir sus cambios, permitir la movilidad o inmovilidad de los orgánulos y participar en la división del citoplasma celular (citocinesis) durante los procesos de división celular.

- Los microfilamentos son fibras de unos 7 nm de diámetro, constituidas por dos cadenas de actina. La actina es una proteína globular que puede formar largas cadenas por unión de un gran número de moléculas individuales. Para formar los microfilamentos, dos de estas cadenas constituidas por actina polimerizada se enrollan entre sí formando una espiral. Aparecen especialmente por debajo de la membrana celular, formando una especie de malla que contribuye a mantener la forma de la célula. También son abundantes en las protuberancias celulares (microvellosidades, pseudópodos) o en zonas en las que se dan procesos de comunicación intercelular. Por último, la combinación de la actina con la miosina permite la contracción celular, por ejemplo en las fibras musculares.
- Los filamentos intermedios son ligeramente más gruesos que los microfilamentos. En realidad se usa este nombre para referirse a fibras celulares de diferente composición, que básicamente participan en el mantenimiento de la estructura de la célula, mediante el establecimiento de una red tridimensional que se extiende por toda la célula a modo de un andamiaje interno.
- Los microtúbulos son cilindros huecos de unos 25 nm de diámetro. En un corte transversal, se observa que el cilindro está formado por trece filamentos que, a su vez, son el resultado de la polimerización de una proteína globular, la tubulina que, a su vez, tiene dos subunidades. Los microtúbulos son estructuras muy dinámicas, que pueden formarse o deshacerse según las necesidades de la célula. Su crecimiento tiene lugar en una única dirección, por lo que se dice que están polarizados (uno de sus extremos, el +, es el que crece, mientras que el otro, llamado -, es el lado por el que se van separando las moléculas de tubulina). En general, los microtúbulos están asociados siempre a una estructura llamada centro organizador de microtúbulos. Pueden existir varios de estos orgánulos en la célula, por ejemplo los corpúsculos basales de cilios y flagelos, o los centriolos, pero en todo caso parecen encargarse de dirigir la polimerización y despolimerización de los microtúbulos relacionados con ellos.
La animación anterior muestra un modelo de la polimerización y despolimerización de los microtúbulos.
Las funciones que realizan los microtúbulos son:
- Contribuyen a mantener la forma de la célula.
- Participan en el transporte de partículas y orgánulos por el citoplasma
- Forman parte del huso acromático, que interviene en la división celular de las células animales.
- Forman parte de los cilios y flagelos, y de los centríolos en las células que los presentan.
No hay comentarios:
Publicar un comentario